
Lys, melatonin og våkenhet/søvnighet
Teksten er skrevet av Knut Inge Fostervold (Psykologisk Institutt ved Universitet i Oslo) og Thor Husby (Luminator As).
Med utgangspunkt i beskrivelsen av behov for god belysning som understøtter stabil døgnrytme presenteres resultatene fra et gjennomført FoU-prosjekt. Prosjektet har hatt som mål å søke etter en hensiktsmessig måte å bidra til dette på ved å ta inn en viktig kvalitet fra dagslyset i det elektriske lyset innendørs uten at dette medfører økt energibruk.
PINEA er betegnelsen på en prinsippløsning for formålet som er utviklet i et samarbeid mellom akademiske miljøer og bedriftspartnere. Denne ble benyttet ved gjennomføring av eksperimentet.
Foruten hensynet til energibruk har det vært viktig at den nevnte løsning isolerer biologiske virkninger fra de mer psykologiske. Dette ivaretas ved at det skapes et kjent lysmiljø hvor lysnivå og fargetemperatur i rommet ikke påvirkes i nevneverdig grad.
Forsøket la til grunn at eksponering av biologisk virksomt lys i området 470-480 nm (nanometer) benyttes med en fastsatt energimengde uten variasjoner over en fastsatt tid.
Resultatene fra det eksperiment som beskrives her, viser at den tekniske løsningen virker ved å forskyve produksjon av melatonin på kveldstid. Dette gir grunnlag for å forvente positive effekter for stabilisering av døgnrytme ved bruk gjennom dagen i tråd med annen nyere forskning. På dette grunnlag planlegges nye studier hvor grupper av brukere følges over lengre tid.
Bakgrunn
Betydningen av god belysning har lenge opptatt både forskere og praktikere. Dette har blitt stadig viktigere etter hvert som overgangen fra industriproduksjon til kunnskapsbasert økonomi har tiltatt. Vi oppholder oss svært mange timer innendørs både på arbeid og hjemme hver dag i kunstig lys.
På forskningssiden er det økende vitenskapelig støtte for antagelsen om at dårlig tilrettelegging av det fysiske arbeidsmiljøet, inkludert belysningen, har betydelige omkostninger. Den canadiske forskeren Vischer (2008) understreker viktigheten av basisbehovene hver enkelt av oss har til miljøfaktorer som lys, luft, renhold og sikkerhet. Dette er faktorer som må være sikret på enhver arbeidsplass og som søkes ivaretatt gjennom lover, forskrifter og måletekniske standarder.
Nyere forskning peker på at effekten av god belysning er mer omfattende enn tidligere antatt. Oppdagelsen av ikke-visuelle ganglion celler i øyet (ipRGC) viser at lys, i tillegg til å gi oss muligheten til å se, også har betydelige ikke-visuelle effekter på mennesket (van Bommel, 2006). Spesielt har man vært opptatt av sammenhengen mellom lys, opplevelsen av våkenhet og konsentrasjon. Hormonet melatonin har en sentral plass i denne diskusjonen. Hormonet produseres i epifysen (glandula pinealis), og det er velkjent at melatonin har stor betydning for vår døgnrytme.
Døgnrytmen styres av en biologisk klokke som hos mennesker er lokalisert til en gruppe nerveceller i hypothalamus i hjernen. Denne cellegruppen går ofte under betegnelsen nucleus suprachiasmaticus (SCN).
Døgnrytmeforstyrrelser er kjennetegnet av et misforhold mellom den indre døgnrytmen og det ytre sosiale og fysiske miljøet (Pallesen og Bjorvatn, 2009). Det har også vist seg at den «innebygde klokken» ikke følger et 24-timers døgn. Faktisk er den innebygde døgnrytmen under normale forhold i gjennomsnitt på nærmere 25 timer. Det betyr at den biologiske klokken må justeres eller nullstilles hver dag (Bjorvatn, 2018). Døgnrytmeforstyrrelser leder ofte til søvnighet, søvnvansker og redusert søvnkvalitet og er i dag så utbredt at Folkehelseinstituttet har omtalt det som ett av landets mest undervurderte folkehelseproblem. Samtidig ser vi at folk i den industrialiserte del av verden sover mindre enn noen gang. En rapport fra «National Sleep Foundation» i USA rapporterer for eksempel at antallet personer som sover mindre enn 6 timer per natt har økt fra 13 til 19 % i perioden 1999 – 2009 (Welsh et al., 2014). Det er velkjent at søvnmangel påvirker viktige kontrollfunksjoner i hjernen, noe som reduserer vår evne til selvregulering (f.eks., Goel et al., 2009). Dette har igjen betydning for helse og trivsel for den enkelte. Selv om problemene knyttet til søvn er sammensatt, har omfanget av søvnproblemer gitt motivasjon for utvikling av belysning som kan bidra til å minske problemet.
Biologisk virksomt lys forstås her som belysning med effekter som kan måles i kroppen som for eksempel hormonene melatonin og kortisol. Emosjonelle eller psykologiske effekter av evt. endringer i det visuelle miljøet, slik de beskrives i publikasjonen fra Licht.wissen (2019) holdes utenfor i denne sammenhengen. Innfallende lys i øyet, spesielt ved bølgelengder i området 470-480 nanometer reduserer og stopper produksjonen av melatonin og bidrar dermed til å nullstille den interne klokken. I tillegg er dette en prosess som er relatert til utskillelsen av kortisol og vår følelse av å være våken og føle seg godt uthvilt (Zisapel, 2006).
Det er godt kjent i forskningslitteraturen at biologisk virksomt lys kan påvirke døgnrytmen og ofte er dette forstått som et resultat av at lys blokkerer melatoninproduksjonen om kvelden. Nyere forskning tyder at lyseksponering i løpet av dagen, hvor det ikke produseres melatonin, også har betydning for helse, søvnkvalitet og døgnrytme. Dette ser ut til å gjelde både for dagslyseksponering (Boubekri, at al., 2014) og biologisk virksomt elektrisk lys (Figueiro et al., 2017; 2019). Ut fra forskningslitteraturen kan det se ut som om vekslingen mellom perioder med lys og mørke har større betydning enn man tidligere har tenkt (Appleman et al., 2013). En forutsetning for dette er imidlertid at lyset har de kvaliteter som er nødvendig for å påvirke biologiske prosesser.
Dette åpner for muligheten til å bruke belysning på en instrumentell måte der man gjennom å etterligne dagslysets spektrale sammensetning kan påvirke og bidra til å stabilisere menneskets døgnrytme. Et problem med denne forskningen er imidlertid at man så langt har benyttet høye lysmengder for å oppnå ønsket effekt (f.eks. Govén, et al., 2011). Dette kan imidlertid være problematisk både i energisammenheng og for arbeids- og oppholdsmiljøet.
Høy belysning trigger de ulike reseptorene i øyet. Ved at ganglionceller med melanopsin foruten å reagere selv også summerer opp signaler fra andre reseptorer, vil høy belysning ha effekt på produksjon av både melatonin og kortisol. Denne artikkelen beskriver imidlertid resultatene av et forsøk med økt lyseksponering i det biologisk mest effektive området på 470-480 nm uten at det medfører økt belysning i rommet. Det stilles krav til at lysmiljøet skal være kjent uten nevneverdige endringer i hverken lysmengde eller fargetemperatur (dvs. ikke ha psykologiske effekter). Energibruken skal ikke påvirkes nevneverdig. Bruk av LED (Light emitting diode) i standard lysarmatur gir nye muligheter for styring av lyset, bl.a. for å blande «riktig type» blått og hvitt lys.
Metode
Design og prosedyre. Studien var et klassisk eksperiment med tid som innengruppe faktor og lysbetingelse som mellomgruppe faktor. Totalt 24 personer i alderen 18 -22 år deltok. Vanlig randomisert tilordning ble brukt til å fordele deltagerne på de to lysbetingelsene. Eksklusjonskriterier var inntak av enkelte matsorter eller bruk av bestemte medikamenter samme dag som eksperimentet ble gjennomført.
I kontrollbetingelsen ble det benyttet lyskilder med samme spektralfordeling som i dagens standard armaturer med 4000K. I eksperimentbetingelsen ble spektralfordelingen endret ved at den armatur som ble benyttet har en tilleggskomponent som gir økt energimengde i området 470-480 nm i hodehøyde i et begrenset område ved pultene. Denne armaturen betegnes PINEA. Tilleggskomponenten i PINEA-armaturen har en relativ intensitet i energimengde i det angitte området som er lik toppunktet i det «blå» området (ved ca. 430-450 nm) for de dioder som inngår i den ordinære armaturen (som ble benyttet i kontrollgruppen). For å gi en pekepinn på intensiteten i den samlede energimengden fra PINEA-armaturen i området rundt 470-480 nm slik det beskrives ovenfor, er det gjort målinger som viser at denne er vesentlig mindre enn 35% av tilsvarende energimengde fra himmellyset i området rundt 470- 480 nm ved sommersolverv på Østlandet i Norge.
Den løsning som ble benyttet gir ikke nevneverdig synlige forskjeller i fargetemperatur i rommet. Total lysmengde i rommet i de to betingelsene var tilnærmet identisk. Samme utenpåliggende nedhengte lysarmatur ble benyttet i begge betingelser. Armaturen var plassert langs forkant på pultene, dvs. delvis over hodet på deltageren, men uten å danne vesentlige slagskygger. Armaturens egenskaper gjør det mulig å isolere de direkte fysiologiske effektene av endret spektralfordeling fra de psykologiske eller innlærte effektene av ulik fargetemperatur, lysstyrke, osv. Alle prosedyrer bortsett fra ulik fordeling i elektromagnetisk stråling var identisk i de to betingelsene.
Design og prosedyre. Studien var et klassisk eksperiment med tid som innengruppe faktor og lysbetingelse som mellomgruppe faktor. Totalt 24 personer i alderen 18 -22 år deltok. Vanlig randomisert tilordning ble brukt til å fordele deltagerne på de to lysbetingelsene. Eksklusjonskriterier var inntak av enkelte matsorter eller bruk av bestemte medikamenter samme dag som eksperimentet ble gjennomført.
I kontrollbetingelsen ble det benyttet lyskilder med samme spektralfordeling som i dagens standard armaturer med 4000K. I eksperimentbetingelsen ble spektralfordelingen endret ved at den armatur som ble benyttet har en tilleggskomponent som gir økt energimengde i området 470-480 nm i hodehøyde i et begrenset område ved pultene. Denne armaturen betegnes PINEA. Tilleggskomponenten i PINEA-armaturen har en relativ intensitet i energimengde i det angitte området som er lik toppunktet i det «blå» området (ved ca. 430-450 nm) for de dioder som inngår i den ordinære armaturen (som ble benyttet i kontrollgruppen). For å gi en pekepinn på intensiteten i den samlede energimengden fra PINEA-armaturen i området rundt 470-480 nm slik det beskrives ovenfor, er det gjort målinger som viser at denne er vesentlig mindre enn 35% av tilsvarende energimengde fra himmellyset i området rundt 470- 480 nm ved sommersolverv på Østlandet i Norge.
Den løsning som ble benyttet gir ikke nevneverdig synlige forskjeller i fargetemperatur i rommet. Total lysmengde i rommet i de to betingelsene var tilnærmet identisk. Samme utenpåliggende nedhengte lysarmatur ble benyttet i begge betingelser. Armaturen var plassert langs forkant på pultene, dvs. delvis over hodet på deltageren, men uten å danne vesentlige slagskygger. Armaturens egenskaper gjør det mulig å isolere de direkte fysiologiske effektene av endret spektralfordeling fra de psykologiske eller innlærte effektene av ulik fargetemperatur, lysstyrke, osv. Alle prosedyrer bortsett fra ulik fordeling i elektromagnetisk stråling var identisk i de to betingelsene.
For å sikre akseptable målenivå av melatonin ble eksperimentet gjennomført på kveldstid. Totalt tok undersøkelsen 4 timer. Ved starten av undersøkelsen ble deltagerne tatt inn i et venterom utelukkende opplyst av stearinlys (20 -30 lux). Prosedyre og informasjon ble gjennomgått og det ble innhentes informert samtykke fra samtlige deltagere. Etter 1 time ble første måling gjennomført og deltageren fordelt til ulike kontor. Belysningsstyrken målt ved Luxmeter på arbeidsfeltene i hvert kontor var justert til et lysnivå som under vanlige betingelser ikke skulle medføre redusert melatoninutskilling (gjennomsnitt: 288 lx i kontrollbetingelsen, 294 lx i eksperimentbetingelsen). Deltagerne fikk ikke benytte noen form for dataskjerm under eksperimentet, men var ellers fri til å benytte tiden som de ville. Forsøksdeltageren oppholder seg på sitt kontor, med samme lyseksponering i 3 timer, fra kl. 21.00 til kl. 24.00. Etter 3 timers eksponering ble siste måling gjennomført og deltagerne dimittert.
Undersøkelsen ble gjennomført i sin helhet i lokalene til Luminator AS. Luminator AS har utviklet og sto for kostnadene til lyskilder og armatur. Salivaprøvene ble analysert av Vitas AS.
Målevariable. Effekten av belysningsbetingelse ble målt ved hjelp av følgende målemetoder.
Bakgrunnsvariabler
Kjønn og alder.
Utfallsvariabler
Melatoninnivå: Melatoninnivå ble målt ved hjelp av salivaprøver. Pretest før lyseksponering ble gjennomført umiddelbart før deltagerne forlot venterommet. Første effektmåling ble gjennomført (Posttest 1) ½ time etter påbegynt eksponering og deretter hver ½ time til 3 timer etter påbegynt lyseksponering. Prøvene ble sentrifugert (10 000 rpm) og fryst umiddelbart etter innsamling. Bühlmann Direct Saliva Melatonin ELISA kit (EK-DSM) (Bühlmann Laboratories AG, Sveits) ble benyttet ved analyse av salivaprøvene. Analysen ble gjennomført i henhold til manualen.
Subjektiv trøtthet: Subjektiv trøtthet ble målt ved hjelp av Karolinska Sleepiness Scale (Gillberg, Kecklund, & Akerstedt, 1994). Pretest måling ble målt rett før deltagerne forlot venterommet. Postest 1 ble gjennomført ½ time etter påbegynt lyseksponering og deretter hver halve time gjennom hele eksperimentet.
Kontrollvariabler
Negativ/positiv affekt: Opplevelse av generell positiv og negativ affekt ble målt 1 gang i løpet av den første halvtimen av lyseksponeringen. Den Norske versjonen av kortversjonen til The positive and negative affect schedule (PANAS) ble benyttet (Thomsen, 2007).
Døgnrytme: Personlig døgnrytme ble målt ved hjelp av den Norske versjonen av: The Circadian Type Inventory – Revised (Milia, Smith, & Folkard, 2005). Målingen ble gjennomført i løpet av den første halvtimen av lyseksponeringen.
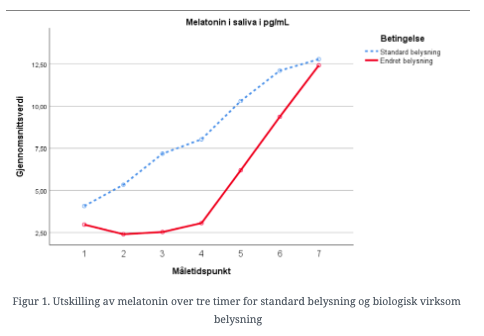
Analysen viser en klart signifikant økning i melatoninnivå for begge grupper i løpet av eksperimentperioden (F(19,1) = 22,50, p < .001). Utviklingen ser imidlertid ut til å være forsinket i eksperimentbetingelsen selv om begge betingelser har omtrent samme nivå ved siste måling. Analysen viser en signifikant kurvelineær forskjell mellom betingelsene (F(19,1) = 6,16, p = .023).
Det var imidlertid ingen særlig forskjell i forløp mellom eksperimentgruppen og kontrollgruppen mht. opplevd søvnighet over tid.
Diskusjon
Formålet med dette FoU prosjektet var å undersøke om endring av spektralfordelingen i området 470 – 480 nanometer påvirker biologiske mekanismer hos mennesker, under normale lysbetingelser (belysningsstyrke og fargetemperatur i rommet samt plassering av armatur). Resultatene viser forsinket utskilling av melatonin under eksperimentbetingelsen sammenlignet med kontrollbetingelsen. Undersøkelsen ser derfor ut til å bekrefte at den tekniske løsningen som er benyttet i PINEA er i stand til å påvirke biologiske ikke-visuelle prosesser. Interessant nok ser det som om denne forsinkelsen kompenseres over tid slik at deltagerne i begge betingelser er omtrent på samme nivå ved siste måling ved midnatt. Dette kan tyde på at påvirkningen ikke er sterk nok til å overstyre fullstendig den biologiske klokken som er med på å bestemme døgnrytmen. Man kan likevel anta at påvirkning over tid vil kunne være med på å forskyve døgnrytmen slik at utskillingen av melatonin starter på et senere tidspunkt.
Det må også opplyses om at intensiteten for energimengden i det biologsk effektive området for påvirkning var satt lavt av forsiktighetshensyn i første forsøk. Intensjonen er at ytterligere forskning skal gi grunnlag for endelig fastsetting av den energimengde som skal benyttes.
Studien finner ingen sammenheng mellom søvnighet og utskilling av melatonin. Hormonet melatonin ser derfor ikke ut til å være knyttet til søvn direkte, men er sannsynligvis en avgjørende faktor for å opprettholde en stabil døgnrytme over tid. Dette funnet er i tråd med tidligere forsking (Rüger, et al., 2005; Nowozin, 2017; Figueiro, 2018). Som nevnt i innledningen er det mye som tyder på at eksponering for biologisk virksomt lys i løpet av dagen har betydning for å opprettholde en stabil døgnrytme. I vår del av verden varierer dagslyset mye i løpet av et år og for mange er det vanskelig å tilrettelegge dagen slik at man kan få utnyttet det naturlige dagslyset. Det er derfor rimelig å anta at økt eksponering for biologisk virksom belysning i løpet av arbeidsdagen kan gi tilsvarende effekter og være med på å stabilisere døgnrytmen. For å undersøke denne hypotesen videre planlegges det studier hvor grupper av brukere følges over lengre tid.
CIE konferanse om biologiske virkninger av lys i Eindhoven (2019)
Artikkelforfatterne deltok på CIE’s konferanse om biologiske virkninger av lys i mars 2019. Konferansen viste at det er stor interesse for feltet i akademiske miljøer i en rekke land og hos næringen.
Flere av de funn som presenteres i denne artikkelen er i tråd med presentasjonene på konferansen, bl.a. forløpet for de to kurvene for melatonin i figuren ovenfor.
Referanser
Appleman, K., Figueiro, M. G., & Rea, M. S. (2013). Controlling light–dark exposure patterns rather than sleep schedules determines circadian phase. Sleep medicine, 14(5), 456-461.
Bjorvatn, B. (2018). Døgnrytmen vår. Nasjonal kompetansetjeneste for søvnsykdommer (SOVno), Lastet ned fra: https://helse-bergen.no/nasjonal-kompetansetjeneste-for-sovnsykdommer-sovno/dognrytmen-var
Boubekri, M., Cheung, I. N., Reid, K. J., Wang, C. H., & Zee, P. C. (2014). Impact of windows and daylight exposure on overall health and sleep quality of office workers: a case-control pilot study. Journal of clinical sleep medicine, 10(06), 603-611.
Figueiro, M. G., Kalsher, M., Steverson, B. C., Heerwagen, J., Kampschroer, K., & Rea, M. S. (2019). Circadian-effective light and its impact on alertness in office workers. Lighting Research & Technology, 51(2), 171-183.
Figueiro, M. G., Steverson, B., Heerwagen, J., Kampschroer, K., Hunter, C. M., Gonzales, K., … & Rea, M. S. (2017). The impact of daytime light exposures on sleep and mood in office workers. Sleep Health, 3(3), 204-215.
Figueiro, M. G., Nagare, R., & Price, L. L. A. (2018). Non-visual effects of light: How to use light to promote circadian entrainment and elicit alertness. Lighting Research & Technology, 50(1), 38-62.
Gillberg, M., Kecklund, G., & Akerstedt, T. (1994). Relations between performance and subjective ratings of sleepiness during a night awake. Sleep, 17, 236-241.
Goel, N., Rao, H., Durmer, J. S., & Dinges, D. F. (2009). Neurocognitive consequences of sleep deprivation. In Seminars in neurology, 29(4), 320-339. Thieme Medical Publishers.
Govén, T., Laike, T., Raynham, P., & Sansal, E. (2011). Influence of ambient light on the performance, mood, endocrine systems and other factors of school children. Proceedings of the 27th Session of the CIE Sun City South Arfica.
Licht.wissen (2019). Guide to Human Centric Lighting (HCL). No 21. Frankfurt am Main: Licht.de. Fördergemeinschaft Gutes Licht.
Nowozin, C., Wahnschaffe, A., Rodenbeck, A., de Zeeuw, J., Hadel, S., Kozakov, R., … & Kunz, D. (2017). Applying melanopic lux to measure biological light effects on melatonin suppression and subjective sleepiness. Current Alzheimer Research, 14(10), 1042-1052.
Milia, L. D., Smith, P. A., & Folkard, S. (2005). A validation of the revised circadian type inventory in a working sample. Personality and Individual Differences, 39, 1293-1305.
Pallesen, S., Bjorvatn, B. (2009). Døgnrytmeforstyrrelser. Tidskriftet Den norske legeforening, 129, 1884-7.
Rüger, M., Gordijn, M. C., Beersma, D. G., de Vries, B., & Daan, S. (2005). Weak relationships between suppression of melatonin and suppression of sleepiness/fatigue in response to light exposure. Journal of sleep research, 14(3), 221-227.
Thompson, E. R. (2007). Development and Validation of an Internationally Reliable Short-Form of the Positive and Negative Affect Schedule (PANAS). Journal of Cross-Cultural Psychology, 38(2), 227-242. doi:10.1177/0022022106297301
van Bommel, W. J. M. (2006). Non-visual biological effect of lighting and the practical meaning for lighting for work. Applied Ergonomics, 37(4), 461-466.
Vischer, J. C. (2008). Towards an Environmental Psychology of Workspace: How People are Affected by Environments for Work. Architectural Science Review, 51(2), 97-108. doi:10.3763/asre.2008.5114
Welsh, D. T., Ellis, A. P., Christian, M. S., & Mai, K. M. (2014). Building a self-regulatory model of sleep deprivation and deception: The role of caffeine and social influence. Journal of Applied Psychology, 99(6), 1268.
Zisapel, N. (2006). The role of melatonin in sleep regulation. In D. P. Cardenali & S. R. Pandi-Perumal (Eds.), Neuroendocrine correlates of sleep/wakefulness, (pp. 295-310). New York: Springer.
Artikkelen er publisert i Magasinet Lyskultur 03/2019. Magasinet distribueres fire ganger i året til Lyskulturs medlemmer (gratis) og abonnenter.
Abonner her: https://lyskultur.no/produkt/arsabonnement-pa-magasinet-lyskultur/
Siste nummer kan bestilles separat frem til nytt nummer er i salg.



Lyskultur på Light & Building 2022

Dagslys i mørkere arealer i byggene

Case study på OBOS living lab, påvirkning av simulert naturlig lys



Duckpin – Europas første kortbane bowling

Dynamisk belysning gir enorme energibesparelser

Den første lysdesigneren

Begynner i jobben som daglig leder i Lyskultur i januar

Ny PhD i dagslys og belysning fra Light and Color centre NTNU

Lysstoffrør fases ut i 2023
